



Наша родословная: Гоминиды (Часть 4)
Уважаемые пикабушники, представляем Вашему вниманию Третью часть серии постов "Наша родословная" (Первая часть, Вторая часть, Третья часть)
Парантроп Бойса (Paranthropus boisei)
Время существования 2.5 млн.л.н. - 1.0 млн.л.н.
Кем впервые описан: Лики Мэри
Когда описан: 1959
Местонахождения: Кения, Танзания

На иллюстрации: Paranthropus boisei. Реконструкция выполнена Романом Евсеевым.
У этих австралопитеков максимального развития достиг челюстной аппарат. Первый открытый череп этого вида из-за размеров зубов даже получил прозвище "Щелкунчик".
Череп P. boisei хорошо известен по множеству находок хорошей сохранности, с датировками в пределах 1,7-1,9 млн.л. Эти черепа очень похожи, их небольшие отличия могут быть отнесены за счет возраста и пола. Череп массивный, с хорошо развитым мускульным рельефом. Конфигурация мозговой коробки своеобразная, при взгляде сверху напоминает кувшин: спереди череп очень расширен благодаря мощному развитию лицевой области, далее следует сильное позадиглазничное сужение, лоб сильно скошен и сужен, потом черепная коробка резко расширяется. Огромные скуловые дуги служат мощными укрепляющими элементами, благодаря своей винтообразной изогнутости. В затылочной норме череп невысокий и широкий в нижней части.
Надглазничный рельеф P. boisei развит достаточно сильно. Размер сагиттального гребня, как и у понгид, различается у особей разного пола: большой у самцов и низкий у самок.
Лицевой скелет P. boisei, как и мозговой, очень своеобразный. Бросается в глаза чрезвычайная расширенность лица, обусловленная огромными размерами верхней челюсти и скуловой кости. Средняя лицевая и скуловая ширина намного превышают как остальные широтные, так и высотные размеры.

На иллюстрации: Paranthropus boisei. Реконструкция выполнена Романом Евсеевым.
Строение мозга P. boisei довольно примитивное. Эндокраны характеризуются сильно суженной лобной долей даже по сравнению с более древними грацильными австралопитеками.
Верхняя челюсть P. boisei очень большая и массивная, очень высокая. Нижняя челюсть, как и верхняя, отличается огромными размерами и массивностью. Относительные размеры передних и заклыковых зубов P. boisei резко отличаются от пропорций, характерных для грацильных австралопитеков. Это классический вариант заклыковой мегадонтии, когда клыки и резцы редуцированы, а премоляры и моляры, напротив, гипертрофированы в размерах. При этом размер моляров увеличивается от первого к третьему. Эмаль зубов очень толстая, возможно, пища массивных австралопитеков состояла из грубоволокнистой, жесткой растительности.
Всё строение черепа P. boisei говорит о больших нагрузках, приходившихся на него; это выражается в большой толщине черепных стенок, развитии рельефа, общей архитектуре с массивными укрепляющими элементами, гипертрофированных челюстях. Жевательный аппарат P. boisei был чрезвычайно развит, все развитие костного рельефа и морфология нижнечелюстного сустава говорят о большой мощности жевательных мышц. Очевидно, массивные австралопитеки были адаптированы к питанию растительной пищей, причем, судя по всему, количество этой пищи было весьма значительным.
Известные находки:
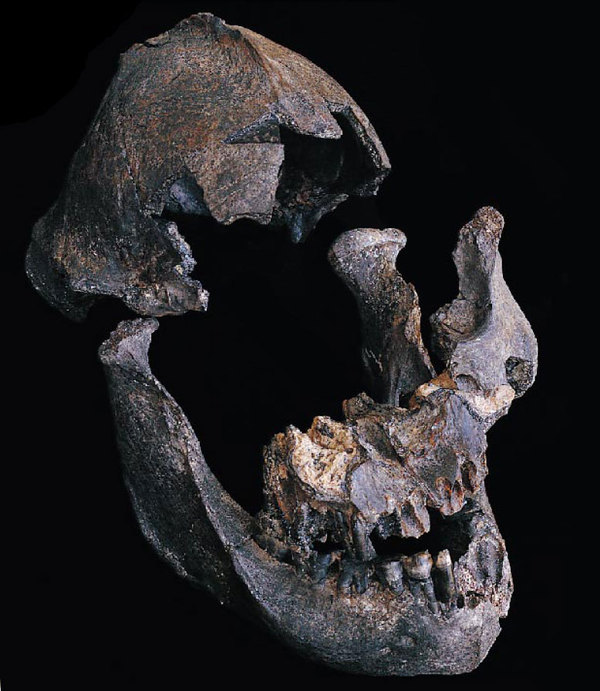
На фото: Нижняя челюсть KNM Peninj 160.

На фото: Деформированный череп KNM-CH 1.
А также:
• Фрагментарный череп KNM-ER 13750
• Нижняя челюсть подростка, с зубами KNM-ER 1477a
• Фрагментарный череп KNM-ER 23000
• Нижняя челюсть с зубами KNM-ER 3230
• Фрагменты черепа самки (без лицевого скелета) KNM-ER 407
• Фрагменты черепа самки KNM-ER 732
• Череп молодой особи KNM-WT 17400
• Нижняя челюсть Omo 57-4-1968-41
• Фрагмент нижней челюсти Omo L 74a-21
• Черепная коробка (7-8 лет или 10-12 лет) Omo L338y-6
Парантроп массивный (Paranthropus robustus)
Время существования 2.5 млн.л.н. - 900 тыс.л.н.
Кем впервые описан: Брум Роберт
Когда описан: 1939
Местонахождения: ЮАР

На иллюстрации: Paranthropus robustus. Реконструкция выполнена Романом Евсеевым
В 1938 г. Роберт Брум проводил раскопки в пещерах Стеркфонтейна. 8 июня 1938 г. школьник Герт Тербланш нашел в местечке Кромдрай, расположенном неподалеку от Стеркфонтейна, вмурованный в породу череп. Р. Брум выменял череп у мальчика за 5 плиток шоколада. Череп, а также ряд обнаруженных в этом местонахождении окаменелостей были описаны под новым родовым и видовым названием Paranthropus robustus. Впоследствии в ряде местонахождений Южной Африки были сделаны сотни находок, относящихся к этому виду гоминид.

На иллюстрации: Paranthropus robustus. Реконструкция выполнена Романом Евсеевым
Таким образом Paranthropus robustus является вторым открытым видом австралопитеков.
Череп P. robustus массивный, с толстыми стенками и выраженным рельефом. Лоб низкий, покатый; надбровный валик мощный, надбровные дуги расположены почти горизонтально, отличаясь от покатых у Paranthropus boisei. Лицевой скелет плоский, даже вогнутый. Скулы массивные, высокие.
Верхняя челюсть P. robustus массивная, укороченная. Нижняя челюсть также чрезвычайно массивна. Заклыковые зубы большие, мегадонтные (хотя не в такой степени, как у Paranthropus boisei).
Затылочное отверстие сдвинуто вперед, что является одним из свидетельств прямохождения массивных австралопитеков.
Эндокран P. robustus имеет примитивную форму, мало отличающуюся от формы человекообразных обезьян.
Верхние конечности P. robustus, как и других австралопитеков, представляют собой мозаику понгидных и гоминидных признаков.

На иллюстрации: Paranthropus robustus. (череп DNH 7 ''Эвридика''). Реконструкция выполнена Олегом Осиповым
Морфология тазовых костей P. robustus, несомненно, ближе к человеческой, нежели к обезьяньей. Таз очень широкий и развернутый. Общий комплекс черт говорит о двуногой походке P. robustus, хотя эта походка, возможно, слегка отличалась от походки представителей рода Homo.
Стопа этих массивных австралопитеков, видимо, была достаточно человекоподобной.
В местонахождении Сварткранс, где также обнаружены останки P. robustus, было найдено множество костяных фрагментов со следами стертости на конце. Опытным путем было показано, что аналогичные следы образуются при использовании костей для вылавливания термитов; таким образом, возможно, P. robustus могли использовать костяные орудия.
Известные находки:

На фото: Череп COB 101
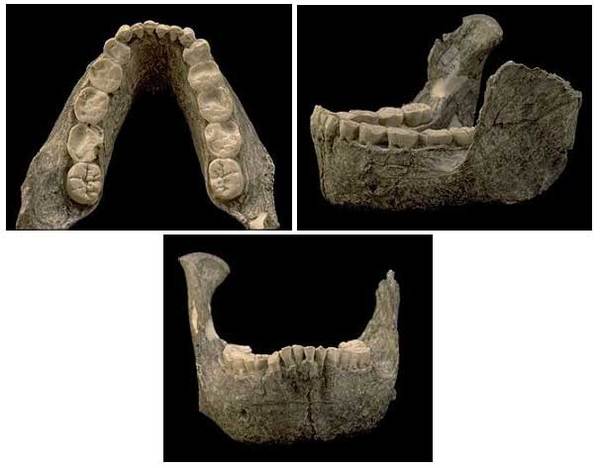
На фото: Нижняя челюсть SK 12.
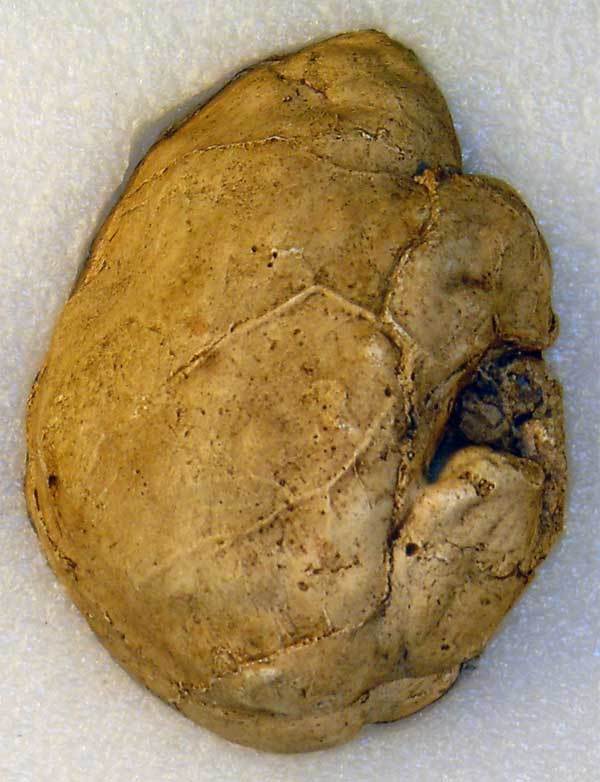
На фото: Естественный слепок мозга SK 1585
А также:
• Основание черепа с верхней челюстью SK 47
• Череп самки 20±1 лет (наиболее полный череп P robustus) SK 48
• Правая тазовая кость взрослой особи SK 50 и SK 3155b
• Лицевая часть черепа с верхней челюстью и фрагментами основания черепа самца 16±1 лет
• Фрагментарный череп, молодого самца SK 54
• Конечная фаланга первого пальца кисти SKX 5016 и др. кости кисти
• Череп с нижней челюстью TM 1517
Человек рудольфийский (Homo rudolfensis)
Время существования 2.3 млн.л.н. - 1.5 млн.л.н.
Кем впервые описан: Алексеев, Валерий Павлович
Когда описан: 1978
Местонахождения: Кения

На иллюстрации: Homo rudolfensis. Реконструкция выполнена Романом Евсеевым
Второй из устоявшихся видов "ранних Homo", сосуществовавший с Homo habilis (т.к. останки обнаружены в тех же местонахождениях и в тех же слоях).
Этот вид «ранних Homo», обладавший крупным мозгом, впервые был описан В.П. Алексеевым как Pithecanthropus rudolfensis в 1978 (в зарубежной литературе всегда фигурирует другой год описания – 1986, являющийся годом выхода перевода книги В.П. Алексеева на английский язык).

На иллюстрации: Homo rudolfensis. Череп Дманиси D4500 с нижней челюстью D2600. Реконструкция выполнена Романом Евсеевым
В последующем взгляд на систематическое положение этого типа гоминид неоднократно пересматривался, наиболее часто используемым стало наименование Homo rudolfensis.
Наиболее характерным отличием от Homo habilis является большая массивность и большие размеры зубов у Homo rudolfensis (хотя некоторые исследователи полагают, что эти отличия могут быть объяснены половым диморфизмом).
Верхняя хронологическая граница существования этого вида довольно расплывчата. Постепенность морфологического перехода между H. habilis и H. rudolfensis, с одной стороны, и первыми архантропами – с другой, не позволяет четко определить момент перехода одного вида в другой.

На иллюстрации: Homo rudolfensis. Череп Дманиси D2700 с нижней челюстью D2735. Реконструкция выполнена Олегом Осиповым
Голотипом вида H. rudolfensis явился широко известный череп KNM-ER 1470, имеющий датировку около 2 млн.л.
Лицо H. rudolfensis заметно уплощенное, а скуловая область развернута вперед. Форма лицевого скелета позволяет сближать H. rudolfensis с более древним Kenyanthropus platyops; некоторые исследователи даже предлагают отнести этот вид «ранних Homo» Кооби Фора к роду Kenyanthropus с образованием названия Kenyanthropus rudolfensis (Collard et Wood, 2001).
Однако в целом H. rudolfensis довольно слабо отличаются от Homo habilis
Для H. rudolfensis характерна умеренно выраженная заклыковая мегадонтия и легкая редукция передних зубов.
Объем мозга H. rudolfensis – более 700 см3 (это гораздо больше, чем у австралопитеков).
См. также материал 2012 г. о новых находках, отнесенных к виду H. rudolfensis.
"Гоша" - трехмерная анимированная реконструкция гоминида из Дманиси (D4500 + D2600).
3D-проект Сергея Кривоплясова специально для портала АНТРОПОГЕНЕЗ.РУ.
Реконструкция подготовлена к открытию выставки "10 черепов, которые потрясли мир"
(Государственный Биологический муеей имени К.А. Тимирязева).
Известные находки:
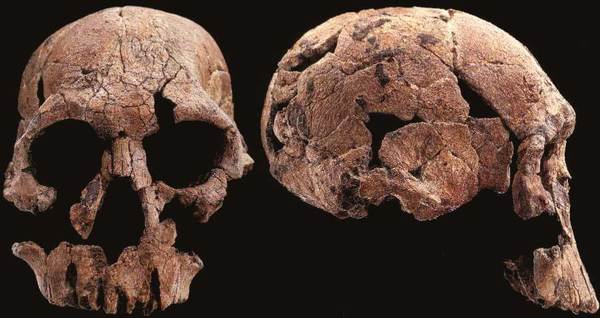
На фото: Череп KNM-ER 1470.


На фото: Бедренные кости KNM-ER 1481(a), KNM-ER 1481(b-d)
А также:
• Нижняя челюсть, изолированные зубы KNM-ER 1482
• Нижние челюсти KNM-ER 1501, KNM-ER 1502, KNM-ER 1801
• Cвод черепа, верхняя челюсть, зубы детёныша 5,2-5,3 лет мужского пола KNM-ER 1590 a-q
• Нижняя челюсть мужчины KNM-ER 1802
• Черепная крышка, левая скуловая кость; натуральный эндокран KNM-ER 3732
• Фрагменты черепа KNM-ER 3891
• Нижняя челюсть KNM-ER 60000 и фрагмент нижней челюсти KNM-ER 62003
• Нижняя челюсть с зубами, часть лица подростка ~8 лет KNM-ER 62000
• Фрагменты черепа и нижней челюсти Omo 75-14
• Тело нижней челюсти Omo 75-146
• Тело нижней челюсти с P1, P2, M1, M2 мужчины UR-501
• Черепная коробка молодого мужчины Дманиси 2280
• Череп девушки 15-16 лет Дманиси 2282
• D-2600, Homo georgicus Дманиси 2600
• D-2700 + D-2735 = Skull C Дманиси 2700 + 2735
• D-3444 + D-3900 Дманиси 3444 + 3900
• Части скелетов подростка (череп Дманиси 2700) и еще трех индивидов.
Время существования 2.3 млн.л.н. - 1.5 млн.л.н.
Кем впервые описан: Нэйпир Джон Рассел, Тайеб Морис, Лики Мэри, Лики Льюис
Когда описан: 1964
Местонахождения: Кения, ЮАР

На иллюстрации: Homo habilis. Реконструкция выполнена Олегом Осиповым
Человек умелый был первым из наших предков, который стал регулярно изготавливать каменные орудия труда и перешёл на всеядность; с человека умелого начинается бурный рост размеров головного мозга. Homo habilis был создателем ранней галечной (так называемой «олдувайской» или «олдованской») культуры.
Homo habilis иногда определяется как поздний грацильный австралопитек, однако ряд прогрессивных признаков в строении приближает его к более поздним гоминидам.

На иллюстрации: Homo habilis. (Череп KNM-ER 1813) Реконструкция выполнена Олегом Осиповым
Череп Homo habilis хорошо известен по множеству находок хорошей сохранности. Череп во многом схож с австралопитеками, однако обладал более округлым и высоким сводом, а мозговая полость - имела в среднем более крупные размеры; костный рельеф черепа был довольно слабо развит. Боковые стенки черепа H. habilis почти вертикальные, относительно высокие. Лобная кость убегающая назад, однако выпуклая в большей степени, чем у австралопитеков; надбровье мощное, выступающее. Затылочная кость высокая, округлая, рельеф ее очень слабый, у H. habilis нет характерных для австралопитеков затылочных гребней. Затылочное отверстие вытянутое, смещено вперед на основании черепа, что свидетельствует о прямохождении.
Лицевой скелет H. habilis более прогрессивный, чем у грацильных австралопитеков. Горизонтальная уплощенность лица в большинстве случаев слабая, носовая область выступает вперед, тогда как скулы сдвинуты назад.

На иллюстрации: Homo habilis. Реконструкция выполнена Романом Евсеевым
Челюсти H. habilis имеют меньшие размеры и выступают слабее, нежели у австралопитеков.
Зубы H. habilis в среднем меньше, чем у грацильных и массивных австралопитеков, но превосходят размерами зубы более поздних архантропов. Таким образом, отчетливо проявляется эволюционная тенденция к общему постепенному уменьшению размеров зубов.
Пропорциями конечностей H.habilis не отличались от австралопитековых.
Стопа H.habilis была уплощённой, без выраженного свода, но пальцы были прямыми, короткими, а большой палец полностью приведён к остальным.
Известные находки:
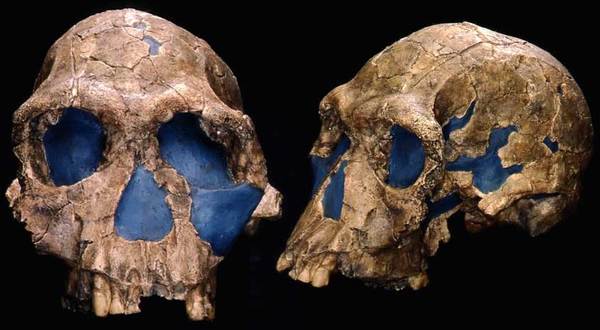
На фото: Череп KNM-ER 1813
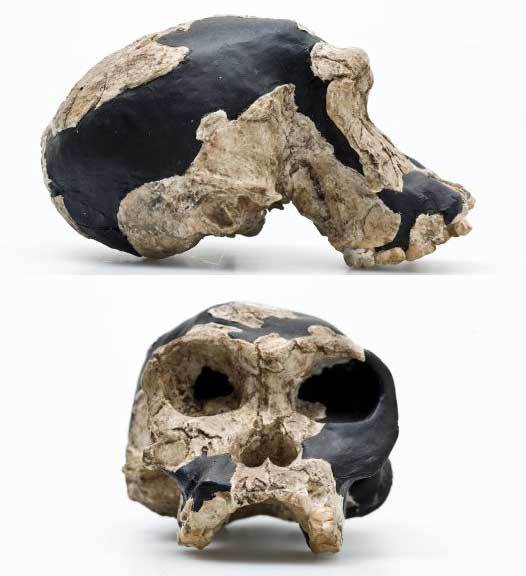
На фото: "Твигги", "Twiggy" Череп OH 24
(К сожалению на этом этапе Пикабу сообщил, что максимально можно добавить 51 блок, так что дальше будет только текст)
А также:
• Верхняя челюсть AL 666-1 из двух половин с нёбом и зубами AL 666-1
• Фрагмент правой височной кости KNM-BC 1
• Бедренные кости: KNM-ER 1472, KNM-ER 1481a
• Нижние челюсти KNM-ER 1501, KNM-ER 1502, KNM-ER 1801
• Фрагмент правой нижней челюсти KNM-ER 1506A
• "Загадочный череп", "The Mystery Skull" KNM-ER 1805
• Фрагментарный скелет: лобная кость, височная кость, кости конечностей взрослого индивида KNM-ER 3735
• Черепная коробка KNM-ER 42700 молодого взрослого
• Левая нижняя челюсть с зубами (клык, премоляры, моляры), взрослый LD 350-1
• Левая большая берцовая и малая берцовая кость OH 35
• Фрагмент левой нижней челюсти OH 37
• Скелет из 302 фрагментов OH 62
• 2 фрагмента правой и левой теменных костей OH 7
• 26 фрагментов черепа и зубы Omo L 894-1
• Череп из 9 фрагментов Stw 53
• Тело нижней челюсти мужчины UR-501
• Фрагмент правого тела нижней челюсти с тремя зубами ребёнка 3/5 лет Гарба IV
Продолжение следует!..
Источник: АНТРОПОГЕНЕЗ.РУ
Показать полностью
24
1
Наша родословная: Гоминиды (Часть 3)
Уважаемые пикабушники, представляем Вашему вниманию Третью часть серии постов "Наша родословная" (Первая часть, Вторая часть)
Приятного чтения!
Австралопитек бахр-эль-газальский (Australopithecus bahrelghazali)
Время существования 3.5 млн.л.н. - 3.0 млн.л.н.
Кем впервые описан: Коппенс Ив, Brunet et al.
Когда описан: 1996
Местонахождения: Чад
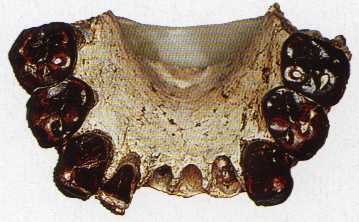
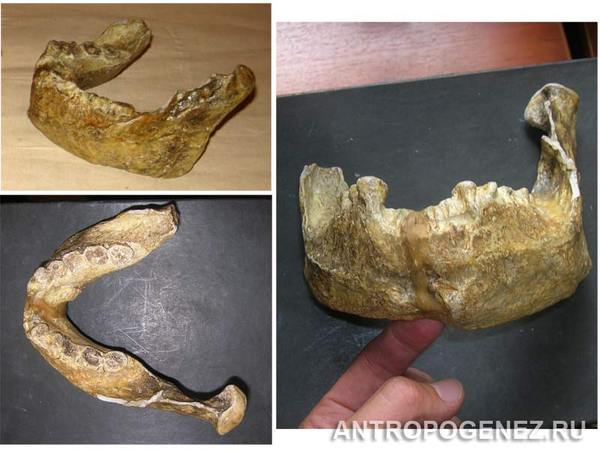
На фото: Передняя часть нижней челюсти с 7 зубами.
Передняя часть нижней челюсти с 7 зубами гоминида была найдена М. Брюнет с сотрудниками и послужила голотипом для описания нового вида Australopithecus bahrelghazali.
Всего обнаружены один фрагмент верхней и один – нижней челюстей, а также изолированный зуб. В целом данные находки мало отличаются от останков афарских австралопитеков.
Своеобразие зубной системы состоит в наличии трех корней у P1 и P2, тогда как у A. afarensis имеется 1-2 корня на P1 и 2 корня на P2; трехкорневые нижние премоляры нехарактерны ни для одного другого вида австралопитековых.
Важность находки заключается в ее географическом положении: большинство австралопитеков найдено в Восточной и Южной Африке, а Австралопитек бахр-эль-газальский - в Северной, что свидетельствует о очень широком распространении группы.
Австралопитек гари (Australopithecus garhi)
Время существования 2.5 млн.л.н.
Кем впервые описан: Лавджой Клод Оуэн, Уайт Тим , Asfaw Berhan et al.
Когда описан: 1999
Местонахождения: Эфиопия

На иллюстрации: Australopithecus garhi. Реконструкция выполнена Олегом Осиповым.
«Garhi» в переводе с афарского означает «удивительный» или «сюрприз».
Это поздний грацильный австралопитек с датировкой около 2,5 млн. лет назад.
A. garhi - eдинственный австралопитек, с останками которого были найдены каменные орудия труда и расколотые кости антилоп; судя по датировке и специализированным чертам, нашим прямым предком не являлся.
Представлен череп, зубы, кости конечностей. Рост 1,2-1,5 м. Объем мозга около 440 см³. Размеры передних зубов A. garhi превосходят таковые у любых других видов австралопитеков.
Известные находки:
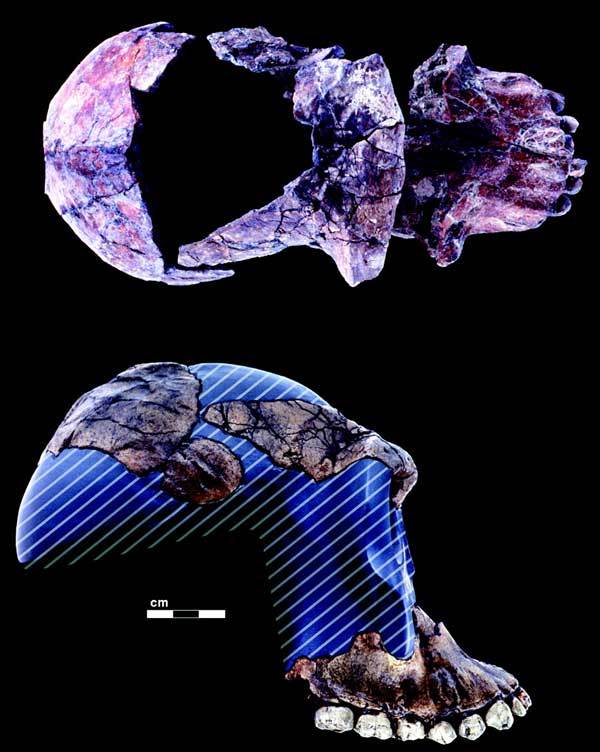
На фото: Фрагменты черепа BOU-VP-12/130.
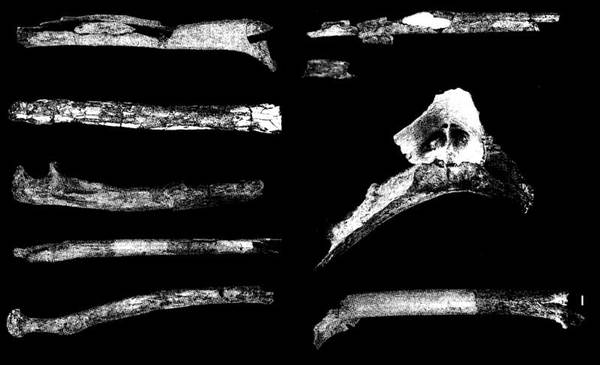
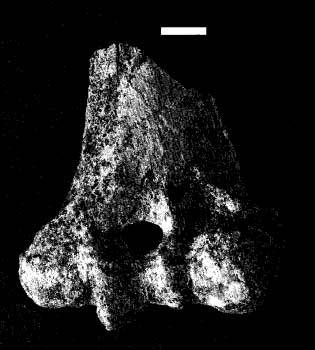
На фото: Дистальный фрагмент левой плечевой кости.
Австралопитек седиба (Australopithecus sediba)
2.0 млн.л.н. - 1.8 млн.л.н.
Кем впервые описан: Berger, Ruiter, Churchill et al.
Когда описан: 2010
Местонахождения: ЮАР

На иллюстрации: Australopithecus sediba. Реконструкция выполнена Романом Евсеевым.
В 2008 году в карстовой пещере, на глубине нескольких десятков метров были обнаружены два разрушенных скелета (взрослой самки и подростка) и отдельные кости третьего (младенца). По этим находкам в 2010 году описан новый вид. Слово "sediba" на местном африканском языке сесото означает «родник».
Интересно, что находка останков Australopithecus sediba сделана с использованием популярного сервиса Google Earth, позволившего выявить местонахождения многочисленных карстовых пещер в Южной Африке.
Подробный анализ находок см. здесь и здесь.
По большинству черт Australopithecus sediba – промежуточен между австралопитеками и Homo (по многим признакам этот вид оказывается даже более гоминидным, чем хабилисы Восточной Африки). Из-за ряда "австралопитековых" черт (таких, как маленький объем мозга, особенности строения рук и ног) данный вид отнесли всё-таки к роду Australopithecus, однако некоторые исследователи не согласны с таким решением, и указывают на черты Australopithecus sediba, сближающие его с Homo - слабо выступающие скулы, форму носовых костей, небольшие коренные зубы, некоторые черты строения мозга, кисти, особенности строения таза и т.д.
Наибольшее сходство у Australopithecus sediba выявляется с Australopithecus africanus, от которого A. sediba, по-видимому, и произошёл. По поводу места A. sediba на "эволюционном дереве" возможны разные версии Во-первых, A. sediba может быть истинным переходным звеном между A. africanus и Homo habilis. Однако, древнейшие "ранние Homo" и в Южной, и в Восточной Африке имеют гораздо большие – как минимум на 300 тысяч лет – датировки; впрочем, эти находки крайне фрагментарны и в будущем, возможно, окажется возможным объединить их с A. sediba в один таксон. Во-вторых, A. sediba может быть представителем собственной южноафриканской линии гоминизации, параллельной восточноафриканской, но чуть более поздней; в связи с этим можно вспомнить A. garhi – ещё одну альтернативную ветвь, дошедшую до уровня изготовления орудий. В-третьих, останки, описанные как A. sediba, могли принадлежать метисам между поздними грацильными австралопитеками и уже сложившимися "ранними Homo", хотя эта версия маловероятна.
Известные находки:
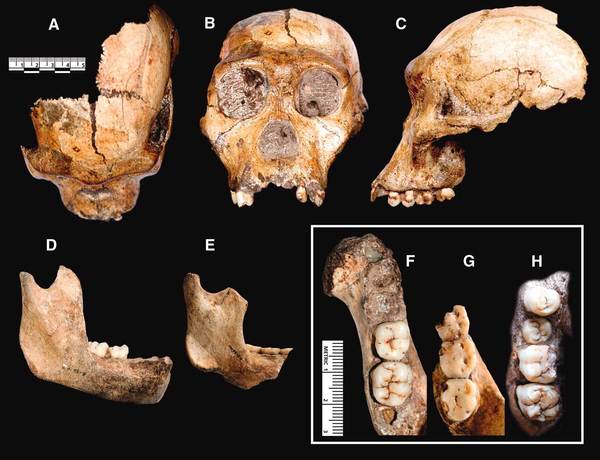
На фото: MH1, MH2: череп и челюсти.
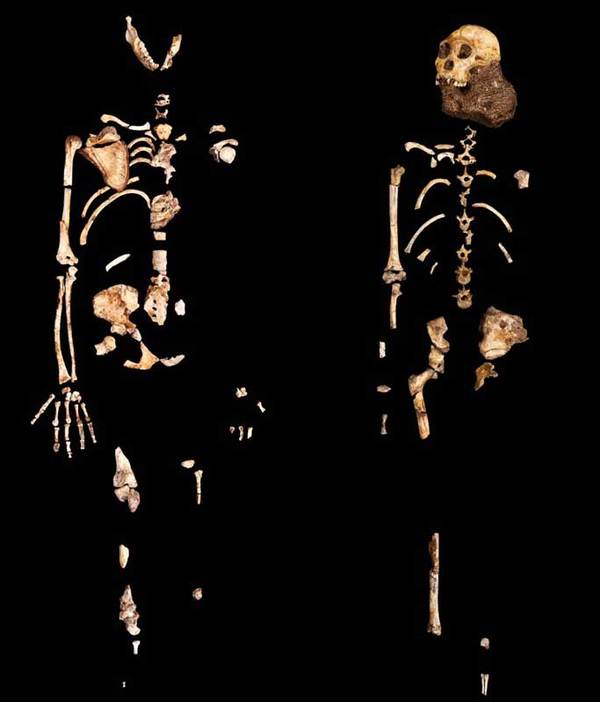
На фото: Скелеты MH2 (слева) и MH1 (справа).
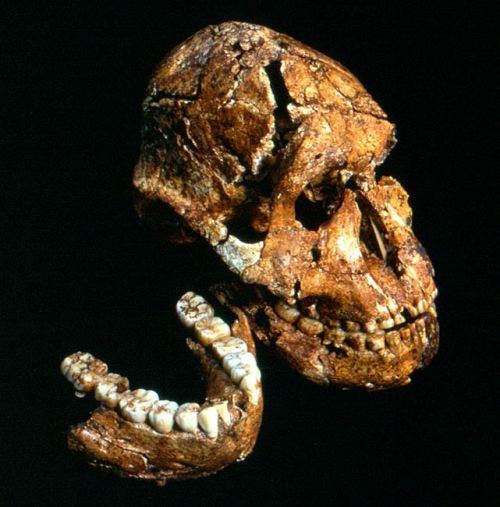
Парантроп эфиопский (Paranthropus aethiopicus)
Время существования 2.5 млн.л.н.
Кем впервые описан: Коппенс Ив, Арамбур К.
Когда описан: 1968
Местонахождения: Кения, Эфиопия

На иллюстрации: Paranthropus aethiopicus. Реконструкция выполнена Романом Евсеевым

На иллюстрации: Paranthropus aethiopicus. Реконструкция выполнена Романом Евсеевым
В 1985 А. Волкером несколько ниже туфа Локалеи был обнаружен почти целый череп KNM-WT 17000, известный также как “Черный череп”, позже были сделаны и другие, менее комплектные находки, например, фрагмент нижней челюсти KNM-WT 16005. Череп KNM-WT 17000 послужил голотипом для описания нового вида.
(Если быть точным, история данного вида довольно запутанная. Первоначально род и вид Paraustralopithecus aethiopicus Arambourg et Coppens, 1968 были описаны на основании морфологии нижней челюсти Omo 18-1967-18 из Омо (2,5 млн.л.н.). Позже, когда был найден череп KNM-WT 17000, он был описан как Australopithecus aethiopicus Walker, Leakey, Harris et Brown, 1986, но прямо не мог быть ассоциирован с Paraustralopithecus aethiopicus Arambourg et Coppens, 1968 (так как там голотип - нижняя челюсть).
Позже этот же череп был описан ещё и как Australopithecus walkeri Ferguson, 1989 (тут была хитрая таксономическая игра с родовыми и видовыми названиями). В итоге договорились называть данный вид Paranthropus aethiopicus, к которому относят и челюсть, и череп, хотя челюсть - всегда с некоторыми оговорками).
Это древнейший из массивных австралопитеков и единственный среди этой группы, имевший резко выступавшие вперёд челюсти.
Череп парантропа эфиопского сочетает в себе примитивные признаки, сближающие его с ранними и грацильными австралопитеками, и специализированные черты массивных австралопитеков.
Paranthropus aethiopicus - вероятный предок более поздних восточно-африканских массивных австралопитеков Paranthropus boisei.
Известные находки:
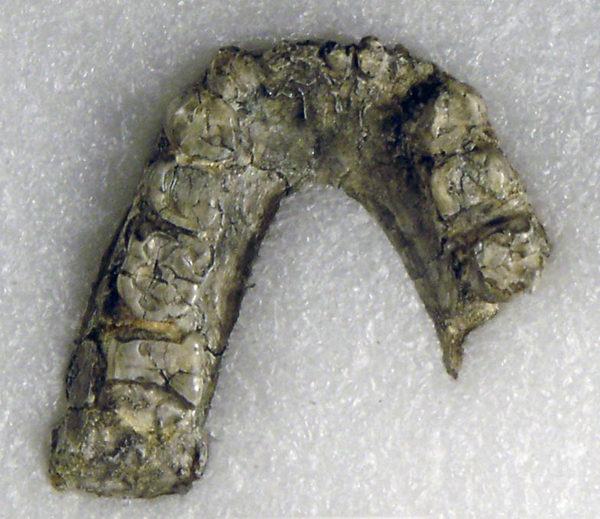

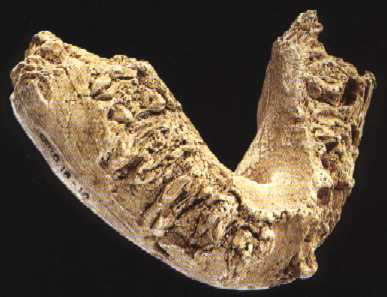
На фото: Нижняя челюсть Omo 18-1967-18.
А также:
• Нижняя челюсть Omo 57-4-1968-41;
• Фрагмент нижней челюсти Omo L 55-s-33;
• Нижняя челюсть Omo L 860-2.
Продолжение следует!..
Источник: АНТРОПОГЕНЕЗ.РУ
Показать полностью
13

Конкурс для мемоделов: с вас мем — с нас приз
Конкурс мемов объявляется открытым!
Выкручивайте остроумие на максимум и придумайте надпись для стикера из шаблонов ниже. Лучшие идеи войдут в стикерпак, а их авторы получат полугодовую подписку на сервис «Пакет».
Кто сделал и отправил мемас на конкурс — молодец! Результаты конкурса мы объявим уже 3 мая, поделимся лучшими шутками по мнению жюри и ссылкой на стикерпак в телеграме. Полные правила конкурса.
А пока предлагаем посмотреть видео, из которых мы сделали шаблоны для мемов. В главной роли Валентин Выгодный и «Пакет» от Х5 — сервис для выгодных покупок в «Пятёрочке» и «Перекрёстке».
Реклама ООО «Корпоративный центр ИКС 5», ИНН: 7728632689
Кому траву, кому крабов: как древние люди делили еду с парантропами
Группа исследователей из университета Барселоны (Испания) обнаружила, что существующие гипотезы о рационе первых представителей рода Homo, а также массивных австралопитеков - парантропов, нуждаются в уточнении. В результате исследования микроскопических царапин на зубах ранних людей и парантропов Восточной Африки учёные пришли к новым выводам, которые представлены в журнале PLoS ONE.
Напомним, что самый древний вид массивных австралопитеков - парантроп эфиопский - жил в Восточной Африке 2,5 млн лет назад. Его вероятным потомком является парантроп Бойса.
На изображении: Paranthropus boisei. Реконструкция выполнена Романом Евсеевым.
Парантропы знамениты своими огромными челюстями и большими жевательными зубами с толстой эмалью. Из-за их внушительного размера первый обнаруженный череп парантропа получил прозвище «Щелкунчик». Эти прямоходящие двуногие обезьяны могли весить до 70 кг. Часто парантропов изображают как ископаемый аналог современных горилл – постоянно жующих вегетарианцев.
Парантропы, судя по всему, длительное время существовали одновременно с ранними людьми – человеком умелым и человеком работающим. Чтобы не мешать друг другу, эти виды обязаны были занимать разные экологические ниши, и их рацион должен был существенно отличаться. Специалисты стараются понять, что же в входило в меню древних гоминид.
Классическая теория заключается в том, что первые люди стали дополнять свой рацион мясом - объедками со стола хищников, а также активно охотиться. Их мозг стал увеличиваться, а челюсти с зубами – уменьшаться. У парантропов всё было по-другому. Считается, что массивные австралопитеки питались растениями cаванн – их меню составляли травы и осоки, а также корневища, клубни, семена и орехи. Для этого и понадобились внушительные челюсти и огромные зубы. Пищевая специализация привела в конечном итоге к вымиранию парантропов. У всеядных людей, которые были менее привередливы в питании, оказалось больше шансов выжить при резких изменениях климата.
Изотопный анализ зубов ранних гоминид подтверждал такие представления. Согласно исследованиям, проводившимся в 2011 и 2013 гг., в рационе восточноафриканских парантропов более 60% составляли растения саванн. У ранних людей саванных растений было меньше, хотя со временем их доля возрастала.
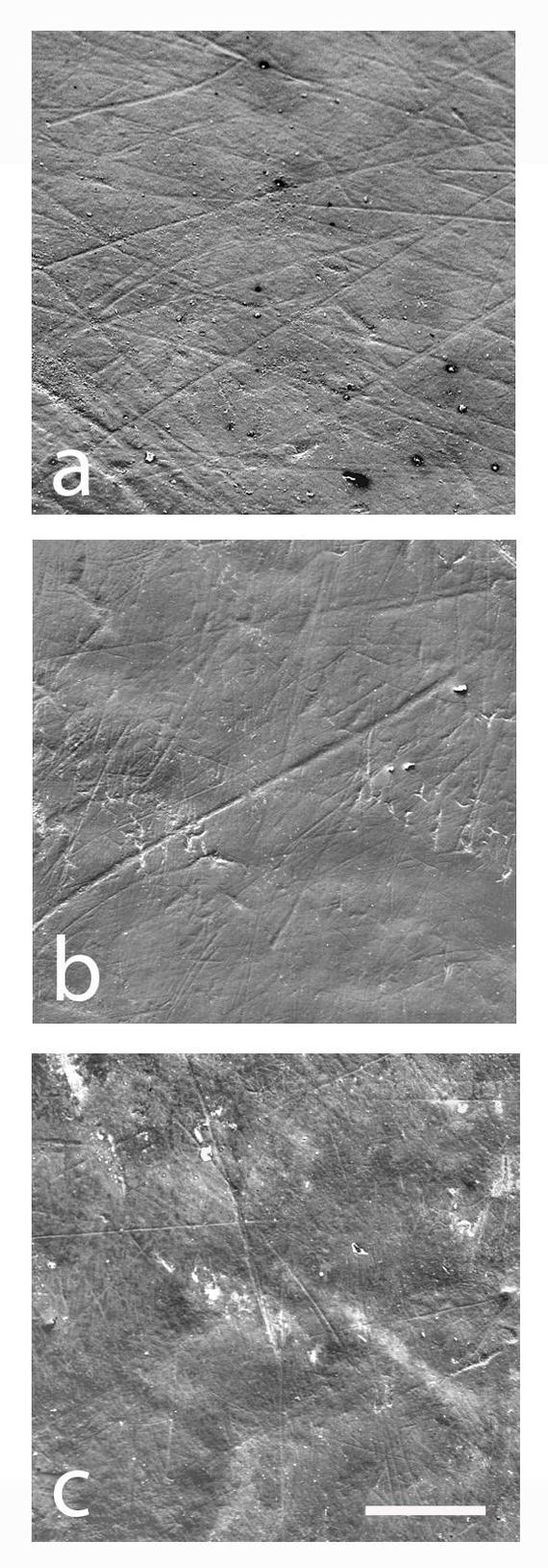
На фото: Щёчная поверхность зубов под электронным микроскопом.
a) LP₄ OH-69 Homo habilis.
b) RM₁ KNM-WT-15000 Homo ergaster.
c) LM₁ Peninj Paranthropus boisei.
Масштаб 200 μm
В 2011 году имеющуюся картину дополнило исследование микроизноса зубной эмали. Дело в том, что в процессе пережевывания пищи на поверхности зубов остаются микроскопические царапины. Их плотность, длина и направленность дают возможность оценить рацион. Если в пище много мелких абразивных частиц – «фитолитов» (кремневых гранул, содержащихся в стеблях травянистых растений) - от их пережевывания на поверхности зубов остается характерный рисунок. Так вот, согласно исследованию микроизноса зубов, парантроп Бойса жевал что-то мягкое, но содержащее эти самые фитолиты. Специалисты предположили, что парантропы питались осокой, растущей вблизи водоемов.
Авторы нового исследования решили использовать еще один источник данных. Если раньше внимание было приковано к жевательной поверхности зуба, то теперь ученые обратили свой взор на «щёчную» сторону зубов. Ведь на этих поверхностях тоже остаются характерные повреждения. Исследования современных животных показали, что износ щечных поверхностей хорошо отражает особенности питания. Кроме того, повреждения на этой стороне зубов накапливаются медленней, чем на быстро обновляющейся жевательной поверхности. Поэтому царапины на щечных поверхностях характеризуют рацион за некоторый более-менее длительный промежуток времени, в то время как жевательная поверхность, фактически, отражает ситуацию на момент смерти животного.
Итак, специалисты изучили микроскопические царапины на 446 зубах, найденных в Эфиопии, Кении и Танзании: 44 зуба парантропа эфиопского, 56 зубов парантропа Бойса, 49 зубов человека умелого (Homo habilis) и 18 человека работающего (Homo ergaster). В лаборатории специалисты сделали с каждого зуба слепок, а затем исследовали его поверхность с помощью растрового электронного микроскопа.
Результаты оказались довольно неожиданными.
Получилось, что ни парантроп эфиопский, ни парантроп Бойса не употребляли жесткой пищи, а ели что-то мягкое и волокнистое. Не было в их пище и значительного количества твердых частиц – фитолитов. Зачем тогда нужны огромные зубы? Возможно, рацион парантропов составляли семена и луковицы? А может быть, парантропы вообще ловили крабов в водоемах (такая гипотеза высказывалась и ранее)? И мощные зубы понадобились для разгрызания панцирей? Однако достаточно ли было подобной пищи в плейстоцене, чтобы прокормить столь крупных едоков? Авторы напоминают, что в Африке царила засуха, озера были соляными и не годились для значительных популяций крабов. Возможно, такая пища была не постоянной, парантропы прибегали к ней лишь эпизодически.
А что можно сказать про рацион ранних людей? Если исходить из полученных данных, хабилисы тоже питались мягкой пищей (мясом?), а у их вероятных потомков – эргастеров рацион становится более разнообразным, и в нем появляется больше твердых продуктов. Авторы предполагают, что это могли быть клубни и корневища. Логично, что более продвинутые Homo пытались использовать всё, что могли добыть в тяжелых климатических условиях (1,8 млн лет назад – время, когда появились первые Homo ergaster – в Африке был пик засухи). Эргастеры могли зависеть от растений не в меньшей степени, чем нынешние охотники-собиратели, типа бушменов. Вероятно – заключают авторы - хабилисы еще тяготели к лесистой местности, где лакомились плодами, а эргастеры окончательно порвали с лесом и освоили саванны.
Как всё это сочетается с данными изотопного анализа? Что ж, авторы просят не забывать о важной детали: стабильные изотопы углерода накапливаются в зубах животного, пока формируется зубная эмаль. То есть в период от нескольких первых месяцев до первых лет жизни. Понимаете, что это значит? Изотопный анализ говорит нам о том, чем питалось данное существо в детстве. Износ зубной эмали, напротив, дает картину на момент смерти или незадолго до нее. Короче говоря, если рацион юного и взрослого животных различался – то изотопы и износ эмали будут давать противоречивые результаты. Вот как всё непросто!
Кстати, мы забыли про южноафриканских парантропов – Paranthropus robustus! В статье про них почти ничего не говорится. Предыдущие исследования показывали, что по своим пищевым пристрастиям эти массивные австралопитеки сильно отличались от восточноафриканских сородичей. Подождем новых результатов?
Источник: АНТРОПОГЕНЕЗ.РУ
Показать полностью
2