В этой статье пойдет речь об удивительном и невероятном, самым примитивно устроенным животным на нашей планете – трихоплаксе (Trichoplax adhaerens, Schultze, 1883).
Trichoplax adhaerens является одним из трех видов в типе Placozoa (Пластинчатые).
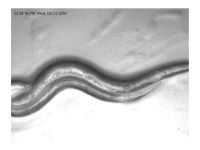
Трихоплакс обычно имеет тонкое уплощенное пластинчатое тело в поперечном сечении около полумиллиметра, иногда до двух или трех миллиметров. Толщина тела обычно составляет всего около 25 мкм. Эти бесцветные серые организмы настолько тонки, что прозрачны, и в большинстве случаев едва различимы невооруженным глазом. Подобно одноклеточным амебам, которым они внешне напоминают, они постоянно меняют свою внешнюю форму (Рис. 1).
Рис. 1. Трихоплакс, размеры около 0,5 мм в диаметре (Автор Bernd Schierwater)
Морфология
У трихоплакса нет тканей и органов; также нет явной симметрии тела, поэтому невозможно отличить переднюю часть от задней или левую от правой. Он состоит из нескольких тысяч клеток шести типов в трех разных слоях:
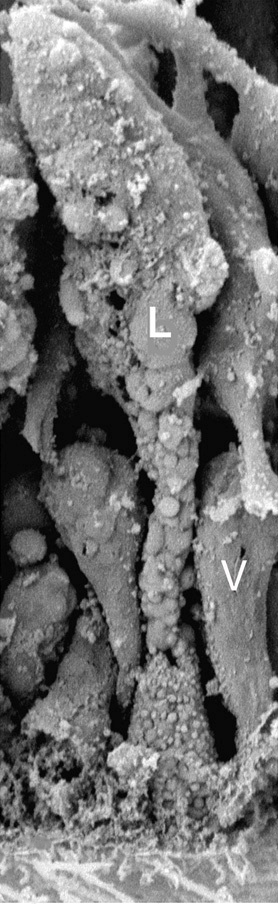
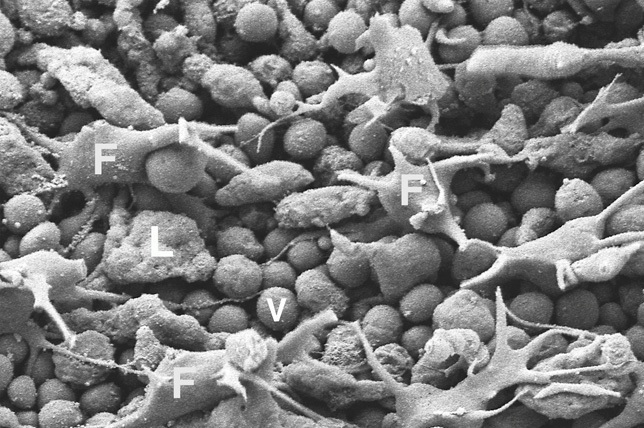
• Вентральные (нижние) эпителиоидные клетки. Мелкие клетки, вытянутые в дорсовентральном направлении, соединены межклеточными контактами и имеют единственную ресничку, поднимающуюся из особого углубления, и множество микроворсинок. На долю этих клеток приходится 72 % всех клеток в теле трихоплакса (Рис. 2).
Рис. 2. Изображение вентральных эпителиоидных клеток (V) и липофильных клеток (L), полученное с помощью электронной микроскопии (Авторы Carolyn L. Smith, Frederique Varoqueau и др.)
• Дорсальные (верхние) эпителиоидные клетки. Подобно вентральным эпителиоидным клеткам, дорсальные клетки соединены межклеточными контактами и имеют реснички и микроворсинки, хотя микроворсинок обычно меньше, чем у клеток вентрального эпителия. Эти клетки составляют 9 % клеток трихоплакса (Рис. 3).
Рис. 3. Изображение дорсальных эпителиоидных клеток (D) и синцитиальных волокнистых клеток (F), полученное с помощью электронной микроскопии (Авторы Carolyn L. Smith, Frederique Varoqueau и др.)
(Почему эпителиоидные, но не эпительные? Потому что эпителий – это ткань, которая должна иметь определенную структуру, клетки в эпителии крепятся к базальной пластинке, а у трихоплакса нет такой штуки. Эпителиоидные – «подобные эпителию»)
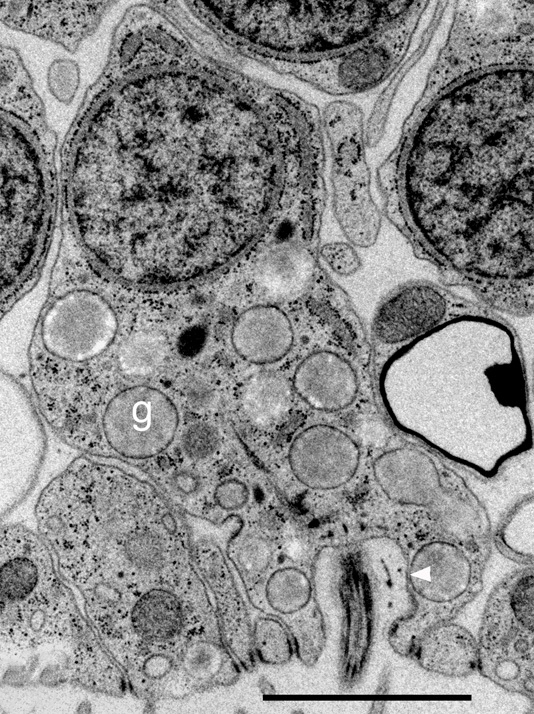
• Железистые клетки. Локализованы среди вентральных эпителиоидных клеток, выполняют секреторную функцию (выделяют слизь на поверхность тела). Железистые клетки наиболее многочисленны в краевой области пластинки (в районе 20 мкм от края), по направлению к центру их численность уменьшается (Рис. 4).
Рис. 4. Микрофотография среза трихоплакса, g - железистые клетки (Авторы Carolyn L. Smith, Frederique Varoqueau и др.)
• Клетки синцитиального волокна. Между двумя слоями клеток (вентральных и дорсальных) находится заполненное жидкостью внутреннее пространство, которое, за исключением непосредственных зон контакта с брюшной и спинной сторонами, пронизано синцитиальным звездчатыми клетками - это волокнистой сетью, состоящей в основном из крупных малочисленных клеток, которые содержат многочисленные ядра, которые, хотя и разделены внутренними поперечными стенками (септами), не имеют настоящих клеточных мембран между ними (Рис. 5).
Рис. 5. Изображение вентральных эпителиоидных клеток (V), липофильных клеток (L) и клеток синцитиального волокна (F), полученное с помощью электронной микроскопии (Авторы Carolyn L. Smith, Frederique Varoqueau и др.)
• Липофильные клетки. Клетки, лишенные ресничек, разбросаны между вентральными эпителиоидными клетками, полностью отсутствуют в дорсальном слое. Участвуют в пищеварении, выделяют крупные гранулы, растворяющие клеточные стенки водорослей. На их долю приходится 11 % клеток в теле трихоплакса, что делает их вторым по многочисленности типом клеток этого организма (Хорошо видны на рисунках 2 и 5).
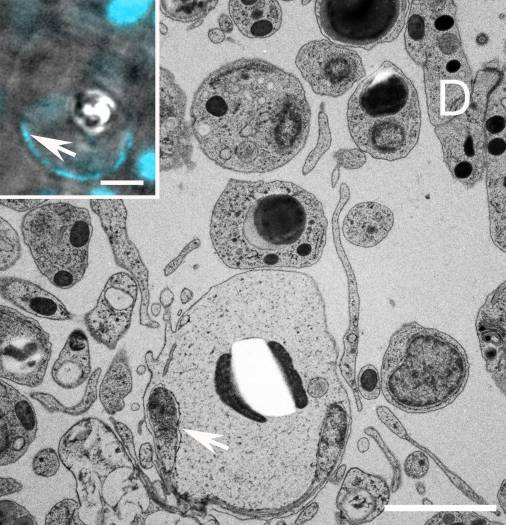
• Кристаллические клетки. Кристаллические клетки содержат ромбоидные кристаллы ~ 2 мкм в диаметре. Они лежат дорсальнее железистых клеток и не имеют входа на поверхность. Точные функции кристаллических клеток неизвестны, но они могут выполнять роль статоцистов или фоторецепторов. Они составляют менее 0,2 % всех клеток в теле трихоплакса (Рис. 6).
Рис. 6. Микрофотография среза трихоплакса, стрелками указаны ядра кристаллических клеток, сами кристаллические включения не могут пережить электронную микроскопию, поэтому на изображении вместо них видно яркое белое пятно, окруженное двумя митохондриями. D - дорсальные эпителиоидные клетки (Авторы Carolyn L. Smith, Frederique Varoqueau и др.)
Общий план строения трихоплакса смотрите на рисунке 7.
Рис. 7. Общий план строения трихоплакса
Обработка сигналов
Как нечто без тканей и органов может воспринимать сигналы окружающего мира, сигналы своих собственных клеток?
В отсутствие нервной системы животное использует короткие пептидные цепочки для связи между клетками, что напоминает то, как животные с нейронами используют нейропептиды для той же цели.
Отдельные клетки содержат и выделяют множество небольших пептидов, состоящих из четырех-двадцати аминокислот, которые обнаруживаются соседними клетками. Каждый пептид можно использовать по отдельности для отправки сигнала другим клеткам, но также последовательно или вместе в разных комбинациях, создавая огромное количество разных типов сигналов.
Это объясняет относительно сложное поведение, такое как сморщивание, повороты, сплющивание и внутреннее вспенивание.
Питание и симбионты
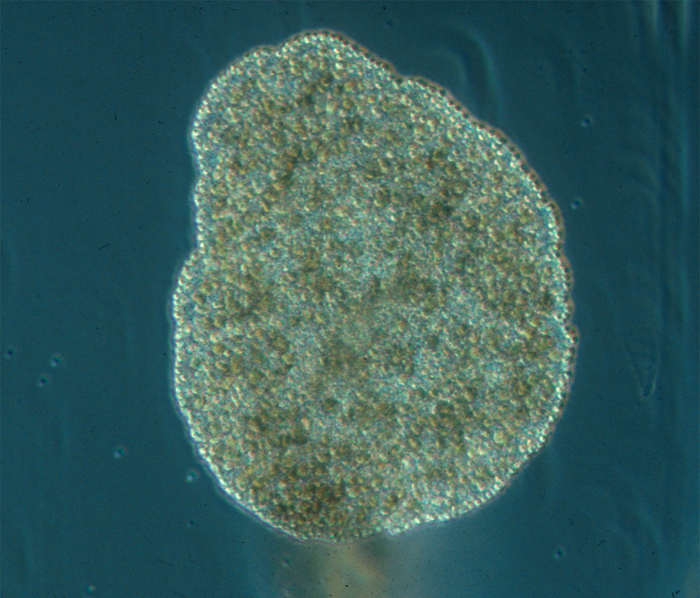
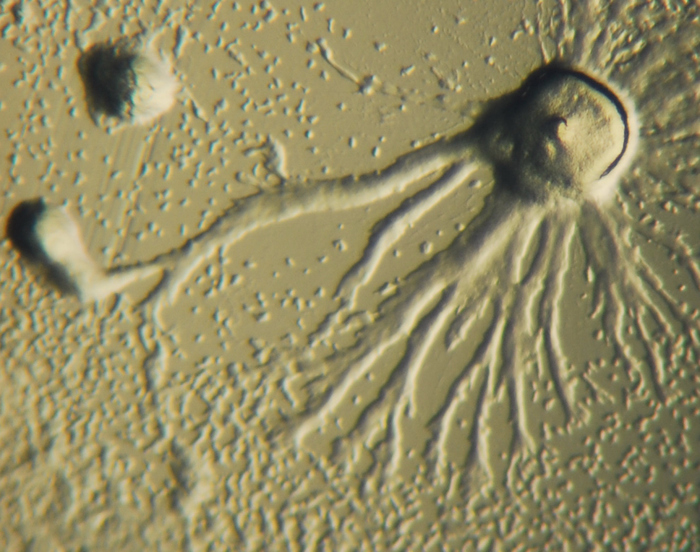
Trichoplax adhaerens питается мелкими водорослями, особенно зелеными водорослями, криптомонадами, а также сине-зелеными бактериями и другими частицами, расположенными на субстрате (детрит) (Рис. 8.)
Рис. 8. Трихоплакс питающийся микроводорослями Rhodamonas salina, t - время в секундах (Автор Carolyn L. Smith)
При питании образуются один или несколько небольших карманов вокруг частиц питательных веществ на брюшной стороне, в эти полости секретируются пищеварительные ферменты; таким образом, организмы развивают временный «внешний желудок», так сказать. Питательные вещества затем поглощаются пиноцитозом («выпиванием клеток») ресничными клетками, расположенными на вентральной поверхности (рис. 9).
Рис. 9. Процесс питания трихоплакса поэтапно (картинка из российской Википедии)
Целые одноклеточные организмы также могут поступать в организм через верхний эпителиоид (то есть «дорсальную поверхность» животного). Этот способ питания может быть уникальным в царстве животных: частицы, собранные в слое слизи, проходят через межклеточные промежутки эпителиоида, а затем фагоцитируются. Такой «сбор» питательных частиц через неповрежденный покров возможен только потому, что некоторые «изолирующие» элементы (в частности, базальная пластинка под эпителиоидом и некоторые типы клеточно-клеточных соединений) не присутствуют у трихоплакса.
Очень часто в синцитиальных клетках трихоплакса обнаруживаются непереваренные бактерии, такие как риккетсии (хотя это могут быть и паразиты).
На твердых поверхностях трихоплакс может двигаться двумя различными способами:
• покрытая ресничками подошва позволяет трихоплаксу медленно скользить по субстрату;
• они могут изменить местоположение, изменив форму своего тела, как это делает амеба.
Эти движения не являются центрально скоординированными, так как животное не имеет мышечной или нервной ткани (gif 1).
Gif 1. Движения трихоплакса
Удалось установить тесную связь между формой тела и скоростью перемещения трихоплакса:
• При низкой плотности питательных веществ трихоплакс поддерживает постоянную скорость, чтобы находить источники пищи, не теряя времени.
• Как только такой источник идентифицируется по высокой плотности питательных веществ, организм увеличивает свою площадь и тем самым увеличивает поверхность, контактирующую с субстратом. Это увеличивает поверхность, через которую могут поступать питательные вещества. Животное одновременно снижает скорость, чтобы съесть всю доступную пищу.
• Когда переваривание почти завершено, трихоплакс снова уменьшает свою площадь, чтобы двигаться дальше.
Фактическое направление, в котором трихоплакс движется каждый раз, является случайным: если мы измерим, насколько быстро отдельное животное удаляется от произвольной начальной точки, мы находим линейную зависимость между прошедшим временем и среднеквадратичным расстоянием между начальной точкой и текущим местоположением.
Такая взаимосвязь также характерна для случайного броуновского движения молекул, что, таким образом, может служить моделью для передвижения у трихоплакса.
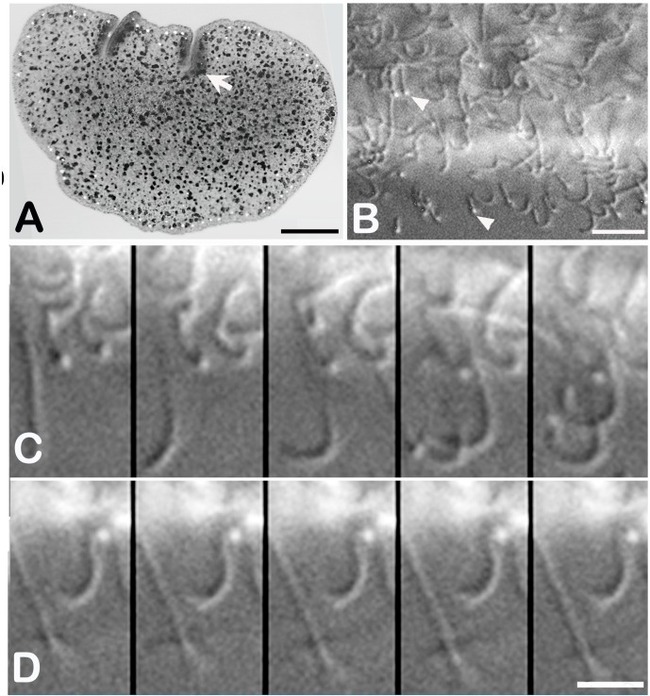
Животные также способны активно плавать с помощью ресничек (Рис. 10). Как только они вступают в контакт с возможным субстратом, возникает дорсовентральная реакция: дорсальные реснички продолжают биться, а реснички вентральных клеток прекращают свое ритмичное биение. В то же время вентральная поверхность пытается войти в контакт с субстратом; небольшие выступы и инвагинации, микроворсинки, обнаруженные на поверхности столбчатых клеток, помогают в прикреплении к субстрату благодаря их адгезивному действию.
Рис. 10. Реснички трихоплакса. А - Трихоплакс на стеклянной подложке с частично поляризованным проходящим светом на конфокальном микроскопе. Инвагинации по верхнему краю (стрелки) - это складки, а темные частицы по всему внутреннему пространству - включения в клетках синцитиального волокна. Яркие частицы вблизи края являются кристаллами в кристаллических клетках. B - Вентральные реснички у края скользящего трихоплакса . Кончики биения ресничек (стрелки) контактируют с подложкой. Животное двигалось к нижней части этого поля. С - пять последовательных изображений (с интервалами 46,5 мс) от животного, показанного на (B), показывают асинхронное биение ресничек. D - вторая последовательность (интервал 46,5 мс) во время паузы после питания, показывающей остановку ресничного биения (Автор Carolyn L. Smith).
Характерной особенностью Placozoa является то, что они могут регенерировать себя из очень маленьких групп клеток. Даже когда большие части организма удаляются в лаборатории, трихоплаксы восстанавливаются в своих размерах. Также можно протирать Trichoplax adhaerens через ситечко таким образом, чтобы отдельные клетки не разрушались, но отделялись друг от друга. В пробирке они снова находят дорогу друг к другу, чтобы сформировать полный организм.
Если эта процедура выполняется на нескольких ранее просеянных через сито животных, то происходит то же самое. В этом случае, однако, клетки, которые ранее принадлежали определенному организму, могут внезапно обнаружиться как часть другого.
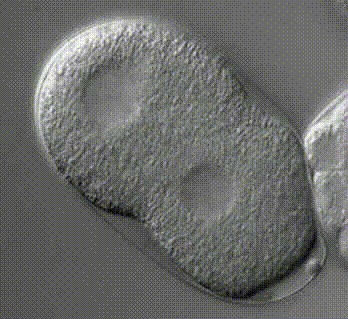
Trichoplax adhaerens размножается преимущественно бесполым путём (фрагментацией или почкованием). При фрагментации тело трихоплакса с помощью образующейся перетяжки разделяется на две примерно равные половинки. Процесс фрагментации может занимать несколько часов, причём между расходящимися особями длительное время сохраняется тонкий многоклеточный мостик (Рис. 11).
Рис. 11. Бесполое размножение трихоплакса (Автор Л.Н. Серавин)
Считается, что половое размножение вызвано чрезмерной плотностью населения. В результате животные впитывают жидкость, начинают набухать и отделяются от субстрата, так что они свободно плавают в воде. В защищенном внутреннем пространстве вентральные клетки образуют яйцеклетку, окруженную специальной оболочкой – мембраной оплодотворения (fertilisation membrane); яйцеклетка снабжается питательными веществами из окружающего синцития, что позволяет накопить богатый энергией желток в его внутренней части.
Как только созревание яйцеклетки завершено, остальная часть животного вырождается, освобождая саму яйцеклетку. Еще не было возможности наблюдать само оплодотворение; существование мембраны оплодотворения в настоящее время считается доказательством того, что это произошло. Предполагаемые яйца наблюдались, но они обычно разлагаются на стадии 32–64 деления. Ни эмбрионального развития, ни сперматозоидов не наблюдалось.
У трихоплакса отсутствует гомолог белка Boule, который, по-видимому, вездесущ и сохраняется у самцов всех протестированных видов других животных. Его отсутствие подразумевает, что у вида нет самцов, и, возможно, его «половое» размножение может быть случаем вышеописанного процесса регенерации, объединяющего клетки, разделенные от двух отдельных организмов в один.
Из-за возможности его клонирования путем бесполого размножения без ограничений продолжительность жизни Placozoa бесконечна; в лаборатории несколько линий, происходящих от одного организма, сохранялись в культуре в среднем 20 лет без возникновения половых процессов.
На этом все про трихоплакса, статья получилась довольно объемной, перелопатил много статей, чтобы все это написать, добавил побольше картинок. Вроде неплохо вышло. Если есть, что добавить, пиши в комменты :)