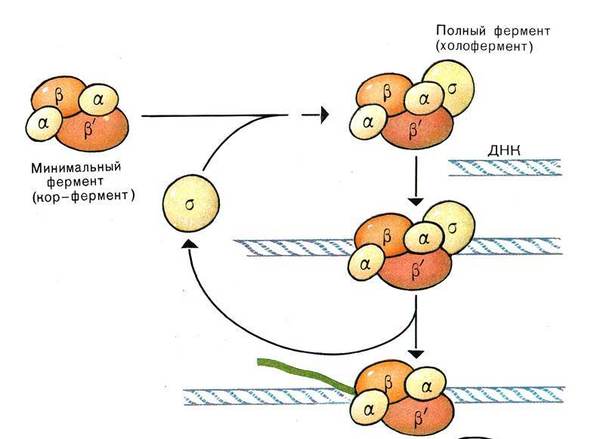
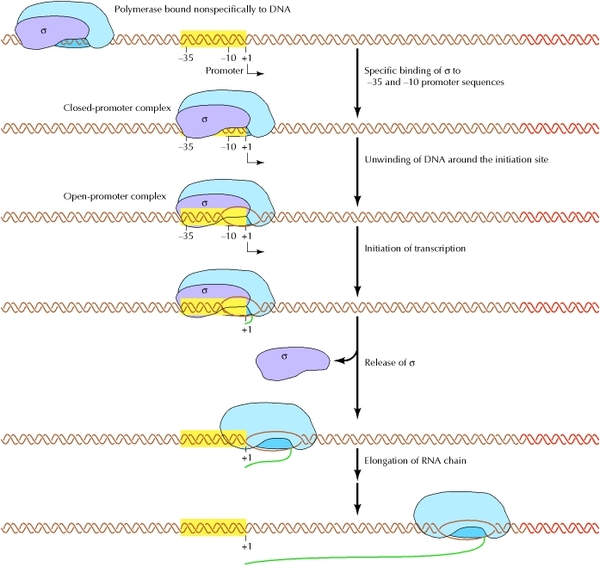
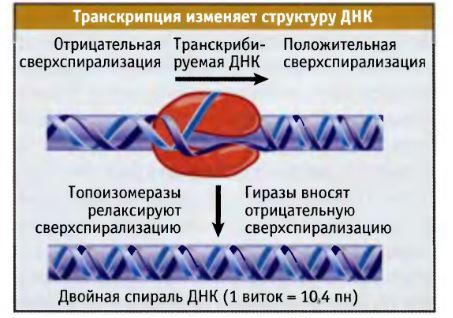
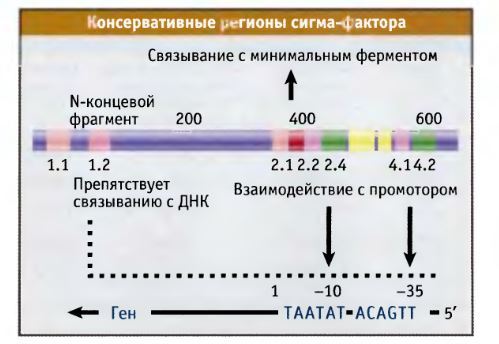
Товарищ @d0nate110 в комментариях к посту попросил сделать цикл постов о работе клетки, тут я поняла, что настал мой час! Пора нести биологию в массы, не будем долго тянуть, Поехали!
Свой рассказ я хочу начать с молекулярной генетики, на мой взгляд самого непонятного и плохо объясняемого раздела молекулярной биологии.
1.Введение
Молекулярная генетика - раздел молекулярной биологии, изучающий процессы связанные с хранением, доступом и реализации генетической информации в клетке.
Вся реализация генетического материала в клетке подчиняется Центральной догме биологии, сформулированной Френсисом Криком (да, тем самым) в 1958 году и дополненной по мере развития молекулярной генетики. Само правило можно выразить схемой:
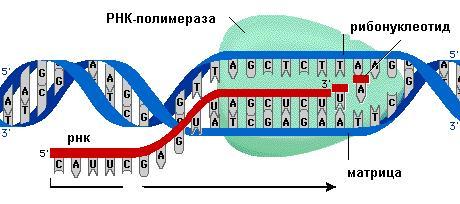
Как видно, что вся генетическая информация хранится в ДНК, люблю приводить аналогию, что ДНК это стоящий дома компьютер с некоторыми файлами. Для дальнейшего использования информации нам стоит переписать ее на более мобильный носитель (флешка), таковой является матричная РНК (мРНК). Переписывание с ДНК на мРНК называется транскрипцией. Далее все эти "знания" нужно использовать, и с мРНК начинает синтезироваться белок. О белках стоит говорить долго и много, поэтому оставим их до следующих постов. Пока нам стоит понять, что белки занимают центральную роль в жизнедеятельности клетки, они разнообразны, как по функциям, так и по строению, состоят из аминокислот. Итак, процесс синтеза белка с мРНК называется трансляцией.
Так в чем же собственно правило? Догма биологии накладывает запрет на "обратную" реализацию информации с белков на ДНК или мРНК, т.е. белок является конечным пунктом в этой всей цепочки исользования информации. Совсем не запрещено совершать информационный переход мРНК-ДНК (обратную транскрипцию), чем и пользуются некоторые хитрые вирусы.
Еще один из важнейших переходов - ДНК-ДНК (репликация). Молекула ДНК, единственная молекула, способная к самокопированию. Это важное свойство нужно для создании полноценной копии всей ДНК клетки в процессе размножения. Репликация обеспечивает точную передачу генетической информации из поколения в поколение.
На схеме не обозначен еще один процесс - репарация ДНК. Пока коротко, репарация - исправление различных повреждений ДНК, которых в клетке совершается огромное количество из-за различных воздействий.
Локальный вывод или что мы должны были узнать из этого раздела:
Существуют 4 процесса реализации генетической информации: (в принципе по каждому из них я бы хотела сделать по небольшому посту)
транскрипция (ДНК-мРНК)
*обратная транскрипция (мРНК-ДНК)
трансляция (мРНК-белок)
репликация (ДНК-ДНК)
и еще один процесс, репарация ДНК - исправление ошибок в последовательности ДНК.
2. Строение ДНК и РНК
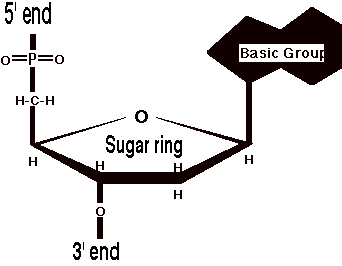
Как мы все уже наверное знаем со школы, ДНК это двойная спираль, но не каждый помнит, что ДНК состоит из нуклеотидов, этаких строительных кирпичиков. Каждый нуклеотид состоит из остатка фосфорной кислоты, присоединённого по 5'-положению к сахару дезоксирибозе, к которому также через гликозидную связь (C—N) по 1'-положению присоединено одно из четырёх азотистых оснований. Если сахар (дезоксирибоза) составляет остов молекулы, то азотистые основания являются именно тем, зачем мы тут и собрались, а именно их последовательность и является той информацией, которая хранится в ДНК.
В состав ДНК входит всего 4 основных азотистых основания! и все наше многообразие зависит от комбинации 4х оснований!!! Вы еще не впечаталились? Тогда идем дальше.
Собственно встречайте 4 основания:
Аденин (А)
Тимин (Т)
Гуанин (Г)
Цитозин (Ц)
Еще одним из важнейших свойств ДНК является комплементарность двух нитей спирали, именно это и помогает держаться им вместе. Комплементарность ДНК - то строгое соответствие соединения азотистых оснований, соединёнными водородными связями, в котором: А-Т (Аденин соединяется с Тимином) Г-Ц (Гуанин соединяется с Цитозином). На этом не сложном правиле основываются все процессы, перечисленные выше в пункте 1.
С молекулой РНК все обстоит немного сложнее. По своей природе РНК очень мобильна и не стабильна (если б вас спросили, где хранить ценную информацию,выбор пал бы на ДНК). Строение РНК очень напоминает ДНК, за исключением 3х пунктов: это одноцепочечная молекула (однако это и помогает ей принимать различные формы, иногда образовывая двуцепочечные фрагменты (все по тому же правилу комплементарности), в состав остова входит рибоза и азотистое основание тимин заменено на урацил (У), т.е. теперь комплементарными становятся А-У. В клетке есть много видов РНК : матричная РНК (мРНК), транспортная РНК (тРНК), рибосомальная (рРНК) и разные малые некодирующие РНК, все они очень занимательны по своим структурам и функциям.
3. Генетический код
Для начала стоит уточнить, что белки состоят из аминокислот, по сути именно аминокислоты и кодируются последовательностью нуклеотидов. Всего биогенными являются 20 аминокислот, т.е. все белки состоят из 20 (!!) аминокислот (а белков у человека по некоторым данным около 70 тысяч!)
Универсальный генетический код - свойственный всем живым организмам (исключения конечно есть, но они не так существенны) способ кодирования последовательности аминокислотных остатков в составе белков при помощи последовательности нуклеотидов в составе нуклеиновой кислоты. Генетический код обладает свойствами:
Триплетность — единицей кода является сочетание трёх нуклеотидов (триплет, или кодон).
Кто-то может уже молодец, и смог посчитать какое количество кодонов может быть составлено из 4х нуклеотидов, если кодон это 3 буквы генетического кода? (64 кодона) Вы спросите, а что так много? А это и есть следущее свойство:
Вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов.
Непрерывность — между кодонами нет знаков препинания, то есть информация считывается непрерывно, будто в предложении нет пробелов.
Неперекрываемость — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов (не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки).
Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте (однако, кодон UGA у Euplotes crassus кодирует две аминокислоты — цистеин и селеноцистеин).
Универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусов до человека (на этом основаны методы генной инженерии; есть ряд исключений, показанный в таблице раздела «Вариации стандартного генетического кода» ниже).
Есть в коде и "знаки препинания": 3 стоп кодона (УАГ, УАА, УГА), старт-кодон, он же АУГ, он же аминокислота метионин, он же старт кодон.
Только для двух аминокислот всего один кодон: метионин, триптофан.
4. Геном и ген (тут мы немного отдохнем)
Геном — совокупность наследственного материала, заключенного в клетке организма. Геном содержит биологическую информацию, необходимую для построения и поддержания жизнедеятельности организма. Большинство геномов, в том числе геном человека и геномы всех остальных клеточных форм жизни, построены из ДНК, однако некоторые вирусы имеют геномы из РНК.
Ген, наверное, самое спорное понятие генетики, никто так и не придумал хорошего и ёмкого определения гену. Тут будем считать, что ген - участок ДНК, с которого происходит транскрипция и в конечном итоге получается продукт.
Каковы же размеры геномов разных видов?
Например, попробуйте решить небольшую задачку:
Дано 6 видов организмов: круглый червь, кишечная палочка, дрожжи, пшеница, мушка дрозофила и человек. Давайте угадаем, у кого же самый длинный геном, а у кого самый маленький, и расположим их в порядке возрастания?
думаем....
думаем...
думаем...
Додумались?
я надеюсь, что вы угадали: кишечная палочка, дрожжи, круглый червь, дрозофила, человек, пшеница.
И правда, обратившись к таблице, мы видим, что не такой уж и человек "венец природы" по длине геномов. И будем сильно заблуждаться, как и заблуждались ученые ранее, считавшие, что чем сложней организм, тем длинней его геном. Сейчас мы рассуждаем в рамках концепции более развитый организм - большее число продуктов (белков), способных получиться из генома, т.е. на передний план выходит соотношение число белков/число генов. (в этом и лидирует). А как получается, что число генов не равно числу белков, а еще и меньше, мы узнаем в следующих постах.
Список литературы по данным темам:
1) Льюин "Гены"
2) Сингер М., Берг П. "Гены и геномы"
3) Ричард Докинз "Эгоистичный ген" (интересный науч поп)
Жду ваши советы, вопросы, пожелания в комментарии.
Следующие посты уже будут более конкретными. Если эта информация оказалась слишком просто и очевидной, не судите строго, тогда сразу нырнем в дебри молекулярной биологии.